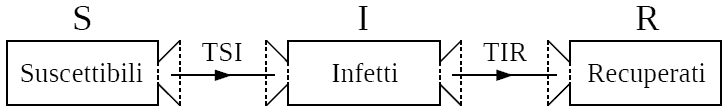
N ≡ popolazione totale
in un intervallo di tempo ∆t.
|
nuovi_infetti | β costante (velocità di contagio unitaria) | |||||
|---|---|---|---|---|---|---|---|
| γI∆t | nuovi_guariti | γ costante (velocità di guarigione unitaria) |
| ∆S | = - nuovi_infetti |
|---|---|
| ∆I | = nuovi_infetti - nuovi_guariti |
| ∆R | = nuovi_guariti |
Sn+1 = Sn + ∆S dato S0
In+1 = In + ∆S dato I0
Rn+1 = Rn + ∆S dato R0
forma discreta |
forma continua |
|||||||||||||||||||||||||||||||||
|
|
nuovi_infetti < nuovi_guariti
βIS/N∆t < γI∆t
β/γ < N/S. Quando accade ?
S<N (Sensibili < Tutti)
matematica: 1<N/S, se β/γ < 1, allora β/γ < N/S
Epidemiologia:
S≈N all'inizio dell'epidemia se prima dell'apparizione dei primi infetti, tutti sono Sensibili all'infezione, poi per un po' rimane S≈N
β/γ < 1 garantisce il diminuire degli infetti da subito.
una questione d'incontri.
L'infezione e' dovuta all'incontro tra suscettibile e infettivo
non tutti gli incontri si concludono con un'infezione.
c∆t nr medio di incontri di 1 persona nella durata ∆t
c tasso di incontro
S/N probabilita' di 1I o di 1R di incontrare S
(S-1)/N probabilita' di un S di incontrare S
per N grande arrotondiamo
S/N probabilita' di 1 S I R di incontrare S
I/N probabilita' di 1 S I R di incontrare I
R/N probabilita' di 1 S I R di incontrare R
c∆tS nr di incontri dei S nella durata ∆t,
I/N e' la frazione di incontri con gli infettivi
p e' la frazione di incontri che trasmette l'infezione
c∆tS*I/N*p nr di nuovi infetti
c∆tI nr di incontri degli I nella durata ∆t,
S/N e' la frazione di incontri con i suscettibili
p e' la frazione di incontri che trasmette l'infezione
c∆tI*S/N*p nr di nuovi infetti
pcSI/N∆t nr di nuovi infetti
β≡pc
βSI/N∆t nr di nuovi infetti
c∆tN nr di incontri nella popolazione di N individui nella durata ∆t.
Gli incontri simmetrici sono trascurabili.
Se tutti incontrassero tutti, gli incontri sarebbero n(n-1)/2, ma non e' il caso, poiche' il nr di incontri di ognuno e' molto minore della popolazione N
es:
allora
solo gli incontri SI sono potenzialmente infettivi
| S | I | R | |
|---|---|---|---|
| S | SS | SI | SR |
| I | IS | II | IR |
| R | RS | RI | RR |
(S+I+R)/N = 1
(S+I+R)/N * (S+I+R)/N = 1*1 = 1
= (SS+SI+SR+IS+II+IR+RS+RI+RR)/N²
pc∆t nr medio di infezioni prodotte da 1 infettivo nella durata ∆t
per calcolare l'evoluzione della popolazione occorre fare il bilancio.
variazione popolazione = (Entrate - Uscite) + (Nascite - Morti)
∆P = (E-U)+(N-M)
| ∆P = E-U | Nascite e Morti = 0 |
| ∆P = -U | solo uscite |
| ∆P = E | solo entrate |
Ipotesi di bilancio del modello SIR: Nascite e Morti = 0
∆S= -US
∆I= EI-UI
∆R= ER
US = EI = TSI
UI = ER = TIR
TSI= βI(S/N)∆t
TIR= γI∆t
β costante, velocità di contagio unitaria
γ costante, velocità di guarigione unitaria
∆I= ∆S - ∆R
It's assumed that the permanence of each single subject in the epidemic states is a random variable with exponential distribution. More complex and realistic distributions (such as Erlang distribution) can be equally used with few modifications.
S+I+R=N = k
∆S+∆I+∆R = 0
∆S= -βI(S/N)∆t
∆R= γI∆t
∆I= ∆S - ∆R
S I R indica la numerosita' delle popolazioni
S(t) I(t) R(t) indica che la numerosita' dipende dal tempo
qui invece di pensare alla sequenza temporale di un individuo durante l'epidemia, si pensa ai gruppi-tipi di popolazione Suscettibili Infetti Recuperati, a come varia lo loro numerosita' in funzione del tempo
Le epidemie si ripetono, sempre con lo stesso andamento, se la popolazione e' abbastanza grande; invece per una popolazione piccola, gli andamenti fluttuano.
c: via alla dimostrazione piu' complicata
∆I < 0
∆I = βIS/N∆t - γI∆t = (βS/N - γ)I∆t
β/γ < N/S ≈ 1 se S≈N cioe' all'inizio dell'epidemia tutti sono Sensibili all'infezione.
β costante, velocità di contagio unitaria
γ costante, velocità di guarigione unitaria